Юрский период климат животные растения. Период геологический. Неогеновый период. Триасовый период. Юрский период. Полезные ископаемые юрского периода
И Швейцарии. Начало юрского периода радиометрическим методом определяется в 185±5 млн. лет, конец — в 132±5 млн. лет; общая продолжительность периода около 53 млн. лет (по данным 1975).
Юрская система в её современном объёме выделена в 1822 немецким учёным А. Гумбольдтом под названием "юрская формация" в горах Юра (Швейцария), Швабский и Франконский Альб (). На территории юрские отложения впервые были установлены немецким геологом Л. Бухом (1840). Первая схема их стратиграфии , расчленения разработана русским геологом К. Ф. Рулье (1845-49) в Подмосковье.
Подразделения . Все основные подразделения юрской системы, впоследствии вошедшие в общую стратиграфическую шкалу, выделены на территории Центральной Европы и Великобритании . Расчленение юрской системы на отделы предложено Л. Бухом (1836). Основы ярусного расчленения юры заложены французским геологом А. д"Орбиньи (1850-52). Немецкий геолог А. Оппель впервые произвел (1856-58) детальное (зональное) подразделение юрских отложений. См. табл.
Большинство зарубежных геологов относит келловейский ярус к среднему отделу, мотивируя это приоритетом трёхчленного деления юры (чёрная, бурая, белая) Л. Буха (1839). Титонский ярус выделяется в отложениях Средиземноморской биогеографической провинции (Оппель, 1865); для северной (бореальной) провинции его эквивалентом является волжский ярус, впервые выделенный в Поволжье (Никитин, 1881).
Общая характеристика . Юрские отложения широко распространены на территории всех континентов и присутствуют в периферии, частях океанских впадин, слагая основание их осадочного слоя. К началу юрского периода в структуре земной коры обособляются два крупных континентальных массива: Лавразия , включавшая в себя платформы и палеозойские складчатые области Северной Америки и Евразии , и Гондвана , объединявшая платформы Южного полушария. Они разделялись Средиземноморским геосинклинальным поясом , который представлял собой океанический бассейн Тетис . Противоположное полушарие Земли занимала впадина Тихого океана , по окраинам которой развивались геосинклинальные области Тихоокеанского геосинклинального пояса .
В океаническом бассейне Тетис в течение всего юрского периода происходило накопление глубоководных кремнистых, глинистых и карбонатных отложений, сопровождавшееся местами проявлениями подводного толеит-базальтового вулканизма. Широкая южная пассивная окраина Тетиса являлась областью накопления мелководных карбонатных отложений. На северной окраине, которая в разных местах и в разное время имела как активный, так и пассивный характер, состав отложений более пёстрый: песчано-глинистые, карбонатные, местами флишевые, иногда с проявлением известково-щелочного вулканизма. Геосинклинальные области Тихоокеанского пояса развивались в режиме активных окраин. В них резко преобладают песчано-глинистые отложения, много кремнистых, очень активно проявлялась вулканическая деятельность. Основная часть Лавразии в ранней и средней юре представляла собой сушу. Морские трансгрессии из геосинклинальных поясов захватывали в ранней юре лишь территории Западной Европы, северную часть Западной Сибири, восточную окраину Сибирской платформы , а в средней юре и южную часть Восточно-Европейской. В начале поздней юры трансгрессия достигла максимума, распространившись на западную часть Североамериканской платформы, восточно-Европейской, всю Западную Сибирь, Предкавказье и Закаспий. Гондвана оставалась сушей на протяжении всего юрского периода. Морские трансгрессии с южной окраины Тетиса захватывали только северо-восточную часть Африканской и северо-западную часть Индостанской платформ . Моря в пределах Лавразии и Гондваны представляли собой обширные, но мелководные эпиконтинентальные бассейны, где накапливались маломощные песчано-глинистые, а в поздней юре в областях, прилегающих к Тетису, — карбонатные и лагунные (гипсо- и соленосные) отложения. На остальной территории юрские отложения либо отсутствуют, либо представлены континентальными песчано-глинистыми, часто угленосными толщами , заполняющими отдельные впадины. Тихий океан в юрском периоде представлял собой типичную океаническую впадину, где накапливались маломощные карбонатно-кремнистые осадки и покровы толеитовых базальтов , сохранившиеся в западной части впадины. В конце средней — начале поздней юры начинается формирование "молодых" океанов; происходит раскрытие Центральной Атлантики, Сомалийской и Северно-Австралийской котловин Индийского океана Амеразийской котловины Северного Ледовитого океана , тем самым начинается процесс расчленения Лавразии и Гондваны и обособления современных материков и платформ.
Конец юрского периода — время проявления позднекиммерийской фазы мезозойской складчатости в геосинклинальных поясах. В Средиземноморском поясе складкообразовательные движения проявлялись местами в начале байоса, в предкелловейское время (Крым, Кавказ), в конце юры (Альпы и др.). Но особенного размаха они достигли в Тихоокеанском поясе: в Кордильерах Северной Америки (невадийская складчатость), и Верхояно-Чукотской области (верхоянская складчатость), где сопровождались внедрением крупных гранитоидных интрузий , и завершили собой геосинклинальное развитие областей.
Органический мир Земли в юрском периоде имел типично мезозойский облик. Среди морских беспозвоночных достигают расцвета головоногие моллюски (аммониты, белемниты), широко распространены двустворчатые и брюхоногие моллюски, шестилучевые кораллы, "неправильные" морские ежи. Среди позвоночных животных в юрском периоде резко преобладают пресмыкающиеся (ящеры), которые достигают гигантских размеров (до 25-30 м) и большого разнообразия. Известны наземные травоядные и хищные ящеры (динозавры), морские плавающие (ихтиозавры, плезиозавры), летающие ящеры (птерозавры). В водных бассейнах широко распространены рыбы, в воздухе в поздней юре появляются первые (зубастые) птицы. Млекопитающие, представленные мелкими, ещё примитивными формами, мало распространены. Растительный покров суши юрского периода отличается максимумом развития голосеменных растений (цикадовые, беннетитовые, гинкговые, хвойные), а также папоротников.
Юрский геологический период, Юра, Юрская система, средний период мезозоя. Начался 200-199 млн л. н. и окончился 144 млн. л. н.
Впервые отложения данного периода были обнаружены и описаны в Юре (горы в Швейцарии и Франции), отсюда и произошло название периода. Отложения юрского периода очень разнообразны: известняки, обломочные породы, сланцы, магматические породы, глины, пески, конгломераты, сформировавшиеся в разнообразнейших условиях. Отложения того времени довольно разнообразны: известняки, обломочные породы, сланцы, магматические породы, глины, пески, конгломераты, сформировавшиеся в разнообразнейших условиях.
Тектоника юрского периода: В начале юрского периода единый суперконтинент Пангея начал распадаться на отдельные континентальные блоки. Между ними образовывались мелководные моря. Интенсивные тектонические движения в конце триасового и в начале юрского периодов способствовали углублению больших заливов, постепенно отделивших Африку и Австралию от Гондваны. Углубился залив между Африкой и Америкой. В Евразии сформировались впадины: Немецкая, Англо-Парижская, Западно-Сибирская. Арктическое море залило северное побережье Лавразии. Именно благодаря этому климат юрского периода и стал более влажным. В юрский период начинают формироваться очертания материков: Африка, Австралия, Антарктида, Северная и Южная америки. И хотя они расположены иначе чем сейчас, но образовались они именно в юрский период.
Климат и растительность юрского периода
Вулканическая активность конца триасового - начала юрского периода вызвала трансгрессию моря. Материки разделились и климат в юрский период стал более влажным чем в триасовый. На месте пустынь триасового периода, в юрский период разрослась буйная растительность. Огромные территории покрылись пышной растительностью. Леса юрского периода в основном состояли из папоротников и голосеменных.
Теплый и влажный климат юрского периода способствовал буйному развитию растительного мира планеты.
Папоротникообразные, хвойные и цикадовые образовывали обширные болотистые леса. На побережье произрастали араукарии, туи, цикадовые. Папоротники и хвощи образовывали обширные лесные массивы. В начале юрского периода, около 195 млн. л. н. на всей территории северного полушария растительность была довольно однообразной. В северном растительном поясе преобладали гинкговые и травянистые папоротники. В юрский период гинкговые были распространены очень широко. Рощи гинкговых деревьев росли по всему поясу.
В южном растительном поясе преобладали цикадовые и древовидные папоротники.
Папоротники юрского периода и в наши дни сохранились в некоторых уголках дикой природы. Хвощи и плауны почти не отличались от современных.
животные: Юрский период - рассвет эпохи динозавров. Именно буйное развитие растительности способствовало появлению множества видов растительноядных динозавров. Рост численности растительноядных динозавров дал толчок к росту численности хищников. Динозавры расселились по всей суше и жили в лесах, озерах, болотах. Диапазон различий между ними настолько велик, что родственные связи между ними устанавливаются с большим трудом. Разнообразие видов динозавров в юрский период было велико. Они могли быть величиной с кошку или курицу, а могли достигать размеров огромных китов.
Юрский период - время обитания многих известных динозавров. Из ящеротазовых это аллозавр и диплодок. Из птицетазовых это стегозавр.
В юрский период в воздухе безраздельно царствовали крылатые ящеры - птерозавры. Они появились еще в триасе, но их расцвет приходился именного на юрский период Птерозавры были представлены двумя группами птеродактили и рамфоринхи.
В юрский период появляются первые птицы или нечто среднее между птицами и ящерами. Существа, появившиеся в юрский период и обладающие свойствами ящеров и современных птиц называются археоптериксы. Первые птицы - археоптериксы, величиной, с голубя. Археоптериксы жили в лесах. Питались преимущественно насекомыми и семенами.
Двустворчатые моллюски вытесняют с мелководий брахиопод. Брахиоподовые ракушечники замещаются устричными. Двустворчатые моллюски заполняют все жизненные ниши морского дна. Многие перестают собирать пищу с грунта и переходят к прокачиванию воды с помощью жабр. В теплых и мелких морях юрского периода происходили и другие важные события.
Юрский период породил множество видов плезиозавров и ихтиозавров, соперничавших с быстроходными акулами и чрезвычайно подвижными костными рыбами. а в морских глубинах леоплеврадон без остановки патрулировал свою территорию в поисках пищи.
Но одно существо по праву можно было назвать хозяином юрских морей. Это гигантский лиоплевродон весом до 25 тонн. Лиоплевродон был самым опасным хищником морей юрского периода, а возможно и за всю историю планеты.
От 213 до 144 млн лет назад.
К началу юрского периода гигантский сверхматерик Пангея находился в процессе активного распада. К югу от экватора все еще существовал единый обширный материк, который снова назвали Гондваной. В дальнейшем он также раскололся на части, образовавшие сегодняшние Австралию, Индию, Африку и Южную Америку. Наземные животные северного полушария уже не могли свободно перемещаться с одного материка на другой, однако они по-прежнему беспрепятственно распространялись по всему южному сверхматерику.
Вначале юрского периода климат на всей Земле был теплым и сухим. Затем, когда обильные дожди начали пропитывать влагой древние триасовые пустыни, мир вновь стал более зеленым, с более пышной растительностью. В юрском ландшафте густо росли хвощи и плауны, которые уцелели с триасового периода. Сохранились и пальмовидные беннеттиты. Кроме того, вокруг было множество гриоов. Обширные леса из семенных, обычных и древовидных папоротников, а также папо-ротникообразных саговников распространились от водоемов в глубь суши. По-прежнему были распространены хвойные леса. Кроме гинкго и араукарий, в них произрастали предки современных кипарисов, сосен и мамонтовых деревьев.
Жизнь в морях.
Когда Пангея начала раскалываться, возникли новые моря и проливы, в которых нашли прибежище новые типы животных и водорослей. Постепенно на морском дне накапливались свежие осадочные отложения. В них обосновалось множество беспозвоночных, таких, как губки и мшанки (морские циновки). В теплых и мелких морях происходили и другие важные события. Там образовались гигантские коралловые рифы, приютившие многочисленных аммонитов и новые разновидности белемнитов (давних родичей нынешних осьминогов и кальмаров).
Па суше, в озерах и реках обитало множество разных видов крокодилов, широко расселившихся по земному шару. Существовали и морские крокодилы с длинными рылами и острыми зубами для ловли рыбы. Некоторые их разновидности даже отрастили ласты вместо ног, чтобы удобнее было плавать. Хвостовые плавники позволяли им развивать в воде большую скорость, чем на суше. Появились и новые виды морских черепах. Эволюция также породила множество видов плезиозавров и ихтиозавров, соперничавших с новыми, быстроходными акулами и чрезвычайно подвижными костными рыбами.

Этот саговник - живое ископаемое. Он почти не отличается от своих родственников, произраставших на Земле в юрский период. Ныне саговники встречаются только в тропиках. Однако 200 млн лет назад они были распространены гораздо шире.
Белемниты, живые снаряды.
Белемниты были близкими родственниками современных каракатиц и кальмаров. Они имели внутренний скелет сигарообразной формы. Его основная часть, состоявшая из известкового вещества, называется ростром. В переднем конце ростра находилась полость с хрупкой многокамерной раковиной, помогавшей животному держаться на плаву. Весь этот скелет размещался внутри мягкого тела животного и служил твердым каркасом, к которому крепились его мышцы.
Твердый ростр лучше всех прочих частей тела белемнита сохраняется в ископаемом виде, и обычно именно он попадает в руки ученых. Но иногда находят и без-ростровые окаменелости. Первые подобные находки в начале XIX в. поставили в тупик многих специалистов. Они догадывались, что имеют дело с останками белемнитов, но без сопутствующего ростра эти останки выглядели довольно-таки странно. Разгадка этой тайны оказалась на редкость простой, лишь только было собрано больше данных о способе питания ихтиозавров - главных врагов белемнитов. По всей видимости, безростровые окаменелости образовывались, когда ихтиозавр, заглотав целый косяк белемнитов, изрыгал мягкие части одного из животных, в то время как его твердый внутренний скелет оставался в желудке хищника
Белемниты, подобно современным осьминогам и кальмарам, вырабатывали чернильную жидкость и использовали се для создания "дымовой завесы", когда пытались спастись от хищников. Ученые обнаружили и окаменевшие чернильные мешки белемнитов (органы, в которых хранился запас чернильной жидкости). Одному из ученых викторианской эпохи, Уильяму Бакланду, удалось даже извлечь из ископаемых чернильных мешков немного чернил, которые он использовал при иллюстрировании своей книги "Бриджуотерский трактат".

Плезиозавры, бочкообразные морские рептилии с четырьмя широкими ластами, которыми они гребли в воде, как веслами.
Склеенная подделка.
Целого ископаемого белемнита (мягкая часть плюс ростр) пока еще никому не удалось найти, хотя в 70-е гг. XX в. в Германии была предпринята довольно-таки изобретательная попытка одурачить весь научный мир с помощью искусной подделки. Цельные окаменелости, якобы добытые п одном из карьеров на юге Германии, несколько музеев приобрели но очень высокой цене, прежде чем обнаружилось, что во всех случаях известковый ростр был аккуратно приклеен к ископаемым мягким частям белемнитов!
Эта знаменитая фотография, сделанная в 1934 г. в Шотландии, была недавно объявлена подделкой. Тем не менее она пятьдесят лет подогревала энтузиазм тех, кто считал лохнесское чудовище живым плезиозавром.
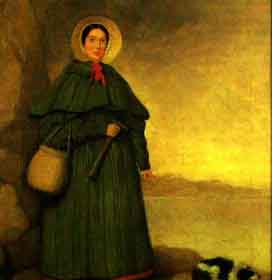
Мэри Эннинг (1799 - 1847) было всего II лет, когда она обнаружила первый ископаемый скелет ихтиозавра у Лайм-Реджис в Дороете, Англия. Впоследствии ей посчастливилось найти также первые ископаемые скелеты плезиозавра и птерозавра.
Ребенок этот мог найти
Очки, булавки, гвозди.
Но тут попались на пути
Ихтиозавра кости.
Рожденные для скорости
Первые ихтиозавры появились еще в триасе. Эти рептилии идеально приспособились к жизни в неглубоких морях юрского периода. Они обладали обтекаемым туловищем, плавниками разной величины и длинными узкими челюстями. Крупнейшие из них достигали в длину примерно 8 м, однако многие виды по размеру не превышали человека. Это были великолепные пловцы, питавшиеся главным образом рыбой, кальмарами и наутилоидеями. Хотя ихтиозавры относились к рептилиям, по их ископаемым останкам можно предположить, что они были живородящими, то есть производили на свет готовое потомство, как млекопитающие. Возможно, детеныши ихтиозавров рождались в открытом море, подобно китам.
Другая группа хищных рептилий, также широко распространенная в юрских морях, - плезиозавры. Их длинношеие разновидности обитали у поверхности моря. Здесь они охотились за косяками нсоольших рыб при помощи своих гибких шей. Короткошеие виды, так называемые плиозавры, предпочитали жизнь па больших глубинах. Они питались аммонитами и прочими моллюсками. Некоторые крупные плиозавры, по всей видимости, охотились также па менее крупных плезиозавров и ихтиозавров.

Ихтиозавры выглядели как точные копии дельфинов, за исключением формы хвоста и лишней пары плавников. Долгое время ученые считали, что у всех ископаемых ихтиозавров, попадавшихся им в руки, был поврежден хвост. В конце концов они догадались, что позвоночник этих животных имел изогнутую форму и на его конце располагался вертикальный хвостовой плавник (в отличие от горизонтальных плавников дельфинов и китов).
Жизнь в юрском воздухе.
В юрском периоде резко ускорилась эволюция насекомых, и в результате юрский ландшафт со временем наполнился нескончаемым жужжанием и потрескиванием, которые издавало множество новых видов насекомых, ползающих и летающих повсюду. Среди них были предшественники
современных муравьев, пчел, уховерток, мух и ос. Позднее, в меловом периоде, произошел новый эволюционный взрыв, когда насекомые начали "налаживать контакты" с только что появившимися цветковыми растениями.
До этого времени настоящие летающие животные встречались лишь среди насекомых, хотя попытки освоить воздушную среду наблюдались и у других существ, научившихся планировать. Теперь же в воздух поднялись целые полчища птерозавров. Это были первые и самые крупные летающие позвоночные животные. Хотя первые птерозавры появились еще в конце триаса, их подлинный "взлет" пришелся именно на юрский период. Легкие скелеты птерозавров состояли из полых костей. У первых птерозавров имелись хвосты и зубы, однако у более высокоразвитых особей эти органы исчезли, что позволило значительно уменьшить собственный вес. У некоторых ископаемых птерозавров угадывается волосяной покров. На основании этого можно предположить, что они были теплокровными.
Ученые до сих пор расходятся во мнениях относительно образа жизни птерозавров. К примеру, первоначально считалось, что птерозавры были своего рода "живыми планерами", парившими, подобно грифам, над землей в потоках поднимающегося горячего воздуха. Возможно, они даже скользили над поверхностью океана, вле-комые морскими ветрами, как современные альбатросы. Однако теперь некоторые специалисты полагают, что птерозавры могли хлопать крыльями, то есть активно летать, подобно птицам. Возможно, одни из них даже ходили по-птичьи, а другие волочили свои тела по земле или спали в местах гнездовий сородичей, повиснув вниз головой, наподобие летучих мышей.

Данные, полученные при анализе окаменевших желудков и помета (копролитов) ихтиозавров, говорят о том, что их рацион состоял в основном из рыбы и головоногих моллюсков (аммонитов, наутилоидей и кальмаров). Содержимое желудков ихтиозавров позволило сделать и еще более любопытное открытие. Маленькие твердые шипы на щупальцах кальмаров и прочих головоногих, судя по всему, доставляли ихтиозаврам немало неудобств, поскольку не переваривались и, соответственно, не могли свободно проходить через их пищеварительную систему. В результате шипы скапливались в желудке, и по ним ученым удается узнать, что данное животное съело на протяжении всей своей жизни. Так, при изучении желудка одного из ископаемых ихтиозавров выяснилось, что он проглотил по меньшей мере 1500 кальмаров!
Как птицы научились летать.
Существуют две основные теории, пытающиеся объяснить, как птицы научились летать. Одна из них утверждает, что первые полеты происходили снизу вверх. Согласно этой теории, все началось с того, что двуногие животные, предшественники птиц, разбегались и подпрыгивали высоко в воздух. Возможно, так они пытались спастись от хищников, а может, ловили насекомых. Постепенно оперенная площадь "крыльев" становилась оолыпе, прыжки, в свою очередь, удлинялись. Птица дольше не касалась земли и оставалась в воздухе. Прибавьте к этому махательные движения крыльями - и вам станет ясно, как спустя длительное время эти "пионеры воздухоплавания" научились подолгу пребывать в полете, а их крылья мало-помалу приобрели свойства, позволявшие им поддерживать тело в воздухе.
Однако существует и другая теория, противоположная, согласно которой первые полеты происходили сверху вниз, с деревьев на землю. Потенциальным "летунам" нужно было сначала забраться на значительную высоту, а уже потом броситься в воздух. В этом случае первым шагом на пути к полетам должно было стать планирование, поскольку при этом типе передвижения энергозатраты крайне незначительны - уж во всяком случае, гораздо меньшие, чем при "бегательно-пры-гательной" теории. Животному не нужно прилагать дополнительных усилий, ибо при планировании его влечет вниз сила земного тяготения.

Первого ископаемого археоптерикса обнаружили спустя два года после опубликования книги Чарлза Дарвина "Происхождение видов". Это важное открытие стало еще одним подтверждением теории Дарвина, гласившей, что эволюция происходит очень медленно и что одна группа животных порождает другую, претерпевая ряд последовательных превращений. Знаменитый ученый и близкий друг Дарвина, Томас Хаксли, предсказал существование в прошлом животного, подобного археоптериксу, еще до того, как его останки попали в руки ученых. По сути дела, Хаксли подробно описал это животное, когда оно еще не было открыто!
Ступенчатый полет.
Один ученый предложил чрезвычайно любопытную теорию. Она описывает ряд стадий, через которые должны были пройти "пионеры воздухоплавания" в ходе эволюционного процесса, превратившего их в конечном итоге в летающих животных. Согласно данной теории, когда-то одна из групп мелких рептилий, именуемых про-топтицами, перешла к древесному образу жизни. Возможно, рептилии забрались на деревья потому, что там было безопаснее, или проще добывать еду, или удобнее прятаться, спать, обустраивать гнезда. На верхушках деревьев было прохладнее, чем на земле, и у этих рептилий для лучшей теплоизоляции развились теплокровность и перьевой покров. Любые лишние длинные перья на конечностях были кстати - ведь они обеспечивали дополнительную теплоизоляцию и увеличивали площадь поверхности крылообразных "рук".
В свою очередь, мягкие, покрытые перьями передние конечности смягчали удар о землю, когда животное теряло равновесие и падало с высокого дерева. Они замедляли падение (выполняя роль парашюта), а также обеспечивали более или менее мягкую посадку, служа естественным амортизатором. Со временем эти животные стали использовать оперенные конечности в качестве протокрыльев. Дальнейший переход от пара-
тютной стадии к планирующей должен был стать вполне закономерным эволюционным шагом, после чего настала очередь последней, полетной, стадии, которой почти наверняка достиг археоптерикс.

"Ранняя" птица
Первые птицы появились на Земле ближе к концу юрского периода. Самая древняя из них, археоптерикс, больше походила на маленького пернатого динозавра, чем на птицу. У нее имелись зубы и длинный костный хвост, украшенный двумя рядами перьев. На каждом ее крыле торчало по три когтистых пальца. Одни ученые полагают, что археоптерикс пользовался своими когтистыми крыльями для лазания по деревьям, откуда он периодически слетал обратно на землю. Другие считают, что он отрывался от земли, используя порывы ветра. В процессе эволюции скелеты птиц становились все легче, а зубастые челюсти заменялись беззубым клювом. У них развилась" широкая грудина, к которой крепились мощные мышцы, необходимые для полета. Все эти изменения позволили усовершенствовать строение тела птицы, придав ему оптимальную для полета структуру.
Первой ископаемой находкой археоптерикса было единственное перо, обнаруженное в 1861 г. Вскоре в той же местности нашли цельный скелет этого животного (причем с перьями!). С тех пор обнаружено шесть окаменевших скелетов археоптерикса: одни полные, а другие лишь фрагментарные. Последняя такая находка датируется 1988 г.
Век динозавров.
Самые первые динозавры появились более 200 млн лет назад. За 140 млн лет своего существования они эволюционировали во множество самых разнообразных видов. Динозавры распространились по всем материкам и приспособились к жизни в самых различных средах обитания, хотя ни один из них не жил в норах, не лазил по деревьям, не летал и не плавал. Одни динозавры были не крупнее белки. Другие весили больше пятнадцати взрослых слонов, вместе взятых. Одни тяжело переваливались на четырех ногах. Другие бегали на двух ногах быстрее, чем олимпийские чемпионы в спринте.
65 млн лет назад псе динозавры внезапно вымерли. Однако перед тем, как исчезнуть с лица нашей планеты, они оставили нам в горных породах подробный "отчет" о своей жизни и своем времени.
Самой распространенной группой динозавров в юрском периоде были прозау-роподы. Некоторые из них развились в самых крупных наземных животных всех времен - зауроподов ("ящероногих"). Это были "жирафы" мира динозавров. Вероятно, они проводили все свое время, поедая листья с верхушек деревьев. Чтобы обеспечить жизненной энергией такое громадное тело, требовалось невероятное количество пищи. Их желудки представляли собой вместительные пищеварительные емкости, непрерывно перерабатывавшие горы растительного корма.
Позднее появилось множество разновидностей небольших быстроногих дино-
завров - так называемых гадрозавров. Это были "газели" мира динозавров. Они щипали низкорослую растительность своими роговыми клювами, а затем пережевывали ее крепкими коренными зубами.
Самым большим семейством крупных плотоядных динозавров были мегалозав-риды, или "огромные ящерицы". Мегало-заврид был чудовищем в тонну весом, с громадными острыми зубами, похожими на зубья пилы, которыми он разрывал плоть своих жертв. Судя по некоторым окаменевшим следам, пальцы его ног были обращены внутрь. Возможно, он передвигался вразвалку, подобно гигантской утке, раскачивая хвост из стороны в сторону. Мегалозавриды заселили все районы земного шара. Их ископаемые останки обнаружены в таких удаленных друг от друга местах, как Северная Америка, Испания и Мадагаскар.
Ранними видами этого семейства были, по всей видимости, относительно небольшие животные хрупкого телосложения. А более поздние мегалозавриды стали поистине двуногими чудовищами. Их задние ноги оканчивались тремя пальцами, вооруженными мощными когтями. Мускулистые передние конечности помогали при охоте на крупных растительно-ядных динозавров. Острые когти, несомненно, оставляли ужасные рваные раны в боку захваченной врасплох жертвы. Могучая мускулистая шея хищника позволяла ему со страшной силой вонзать кинжаловидные клыки глубоко в тело добычи и вырывать из нее громадные куски еще теплого мяса.

В юрском периоде на большей части земной суши разбойничали стаи аллозавров. Они, по всей видимости, представляли собой кошмарное зрелище: ведь каждый член такой стаи весил более тонны. Совместными усилиями аллозавры могли легко одолеть даже крупного зауропода.
Юрский период – это середина мезозойской эры. Этот кусок истории в первую
очередь знаменит своими динозаврами, он был очень хорошим временем для всего
живого. Во время юрского периода впервые рептилии властвовали везде: в воде, на
земле и в воздухе.
Название этот период получил в честь горной цепи в Европе. Начался Юрский период
около 208 миллионов лет назад. Этот период был более революционен, чем
триасовый. Эта революционность заключалась с теми имениями, которые происходили
с земной корой, потому что именно во время юрского периода материк Пангея начал
расходиться. Климат с это времени стал теплее и более влажный. К тому же начал
подниматься уровень воды в мировом океане. Все это дало большие возможности для
животных. Благодаря тому, что климат стал более благоприятный на суше стали
появляться растения. А на мелководьях начали появляться кораллы.
Юрский период длился от 213 до 144 миллионов лет назад. В самом начале Юрского
периода климат на всей Земле был сухим и теплым. Кругом были одни пустыни. Но
позднее их стали пропитывать влагой обильные дожди. И мир стал более зеленым,
стала расцветать пышная растительность.
Папоротникообразные, хвойные и цикадовые образовывали обширные болотистые леса.
На побережье произрастали араукарии, туи, цикадовые. Папоротники и хвощи
образовывали обширные лесные массивы. В начале юрского периода, около 195 млн.
л.н. на всей территории северного полушария растительность была довольно
однообразной. Но уже начиная с середины юрского периода, около 170-165 млн.л.н.,
образовались два (условных) растительных пояса: северный и южный. В северном
растительном поясе преобладали гинкговые и травянистые папоротники. В юрский
период гинкговые были распространены очень широко. Рощи гинкговых деревьев росли
по всему поясу.
В южном растительном поясе преобладали цикадовые и древовидные папоротники.
Папоротники юрского периода и в наши дни сохранились в некоторых уголках дикой
природы. Хвощи и плауны почти не отличались от современных. Места произростания
папоротников и кордаитов юрского периода сейчас занимают тропические леса,
состоящие в основном из сагавников. Саговники - класс голосеменных,
преобладавший в зеленом покрове Земли юрского периода. Ныне они встречаются
кое-где в тропиках и субтропиках. Под сенью этих деревьев бродили динозавры.
Внешне саговники настолько похожи на невысокие (до 10-18 м) пальмы, что даже
изначально были определены как пальмы в системе растений.
В Юрский период так же распространены гинкго - листопадные (что необычно для голосеменных) деревья с кроной, похожей на дуб, и с небольшими веерообразными листьями. До наших дней сохранился только один вид - гинкго двулопастной. Первые кипарисовые и, возможно, еловые появляются именно в юркий период. Хвойные леса юрского периода были похожи на современные.
В юрский период на Земле установился умеренный климат. Даже засушливые зоны были богаты растительностью. Такие условия идеально подходили для размножения динозавров.Среди них выделяют ящеротазовых и птицетазовых.
Ящеротазовые передвигались на четырех ногах, имели на ступнях по пять
пальцев, питались растениями. Большинство из них имели длинную шею, маленькую
голову и длинный хвост. У них было два мозга: один небольшой-в голове; второй
значительно больший по величине – у основания хвоста.
Самым крупным из юрских динозавров был брахиозавр, достигавший в длину 26 м,
весивший около 50 т. У него были столбовидные ноги, маленькая голова, толстая
длинная шея. Жили брахиозавры на берегах юрских озер, питались водной
растительностью. Ежедневно брахиозавру было нужно не менее полутоны зеленой
массы.
Диплодок – древнейшее пресмыкающееся, его длина составляла 28 м. У него была
длинная тонкая шея и длинный толстый хвост. Подобно брахиозавру, диплодок
передвигался на четырех ногах, задние были длиннее передних. Большую часть своей
жизни диплодок проводил на болотах и озерах, где пасся и спасался от хищников.
Бронтозавр был сравнительно высоким, имел большой горб на спине и толстый хвост. Долотовидные маленькие зубы густо располагались на челюстях небольшой головы. Обитал бронтозавр в болотах, на берегах озер. Бронтозавр весил около 30 тонн и превышал 20 и в длину. Ящероногие динозавры (зауроподы) были крупнейшими известными до сих пор наземными животными. Все они были растительноядными. Еще до недавнего времени палеонтологи полагали, что столь тяжеловесные создания вынуждены были проводить большую часть своей жизни в воде. Считалось, что на суше его берцовые кости "подломились бы" под тяжестью колоссальной туши. Однако находки последних лет (в частности, отлечатки ног) говорят о том, что зауроподы предпочитали бродить по мелководью, вступали они и на твердую почву. По отношению к размерам тела, бронтозавры обладали крайне маленьким мозгом, весившим не более фунта. В области крестцовых позвонков бронтозавра имелось расширение спинного мозга. Будучи гораздо более крупным, чем головной мозг, оно управляло мускулатурой задних конечностей и хвоста.
Птицетазовые динозавры подразделяются на двуногих и четвероногих. Разные по величине и внешнему виду, они питались преимущественно растительностью, но среди них появляются и хищники.
К растительноядным относятся стегозавры. Стегозавры особенно обильны в
Северной Америке, откуда известно несколько видов этих животных, достигавших в
длину 6 м. Спина была круто выпуклой, высота животного достигала 2,5 м. Туловище
массивное, хотя стегозавр передвигался на четырех ногах, передние конечности у
него были намного короче задних. На спине в два ряда высились крупные костные
пластины, защищавшие позвоночный столб. На конце короткого, толстого хвоста,
используемого животным для защиты, имелись две пары острых шипов. Стегозавр был
вегетарианцем и имел исключительно маленькую голову и, соответственно, крохотный
мозг, немногим более грецкого ореха. Интересно, что расширение спинного мозга в
крестцовой области, связанное с иннервацией мощных задних конечностей, по
диаметру намного превосходило головной мозг.
Появляется множество чешуйчатых лепидозавров - мелких хищников с клювовидными
челюстями.
В юрский период впервые появляются летающие ящеры. Летали они при помощи кожистой оболочки, натянутой между длинным пальцем кисти и костями предплечья. Летающие ящеры были хорошо приспособлены к полету. Они имели легкие трубкообразные кости. Чрезвычайно удлиненный внешний пятый палец передних конечностей состоял из четырех суставов. Первый палец имел вид маленькой кости либо совсем отсутствовал. Второй, третий и четвертый пальцы состояли из двух, реже трех костей и имели когти. Задние конечности были довольно сильно развиты. На их концах имелись острые когти. Череп летающих ящеров был сравнительно крупным, как правило, удлиненным и заостренным. У старых ящеров черепные кости срастались и черепа становились похожими на черепа птиц. Межчелюстная кость иногда разрасталась в удлиненный беззубый клюв. У зубатых ящеров зубы были простыми и сидели в углублениях. Самые крупные зубы находились спереди. Иногда они торчали в сторону. Это помогало ящерам ловить и удерживать добычу. Позвоночник животных, состоял из 8 шейных, 10-15 спинных, 4-10 крестцовых и 10-40 хвостовых позвонков. Грудная клетка была широкой и имела высокий киль. Лопатки были длинными, тазовые кости срослись. Наиболее характерные представители летающих ящеров – птеродактиль и рамфоринх.
Птеродактили в большинстве случаев были бесхвостыми, разными по величине – от
размеров воробья до вороны. Они имели широкие крылья и вытянутый вперед узкий
череп с небольшим числом зубов в передней части. Птеродактили жили большими
стаями на берегах лагун позднеюрского моря. Днем они охотились, а с наступлением
ночи укрывались на деревьях или в скалах. Кожа птеродактилей была морщинистой и
голой. Питались они главным образом рыбой, иногда морскими лилиями, моллюсками,
насекомыми. Для того, чтобы взлететь, птеродактили вынуждены были спрыгивать со
скал или деревьев.
Рамфоринхи имели длинные хвосты, длинные узкие крылья, большой череп с
многочисленными зубами. Длинные зубы разной величины выгибались вперед. Хвост
ящера заканчивался лопастью, служившей рулем. Рамфоринхи могли взлетать с земли.
Они селились на берегах рек, озер и морей, питались насекомыми и рыбой.
Летающие ящеры жили только в мезозойскую эру, причем их расцвет приходится на
позднеюрский период. Их предками являлись, по-видимому, вымершие древние
пресмыкающиеся псевдозухии. Длиннохвостые формы появились раньше короткохвостых.
В конце юрского периода они вымерли.
Следует заметить, что летающие ящеры не были предками птиц и летучих мышей.
Летающие ящеры, птицы и летучие мыши произошли и развивались каждый своим путем,
и между ними отсутствуют близкие родственные связи. Единственный общий признак
для них – умение летать. И хотя все они приобрели эту способность благодаря
изменению передних конечностей, отличия в строении их крыльев убеждают нас в
том, что у них были совершенно разные предки.
Моря юрского периода населяли дельфинообразные пресмыкающиеся – ихтиозавры. Они имели длинную голову, острые зубы, большие глаза, окруженные костным кольцом. Длина черепа некоторых из них составляла 3 м, а длина тела – 12 м. Конечности ихтиозавров состояли из костных пластинок. Локоть, плюсна, кисть и пальцы по форме мало отличались друг от друга. Около ста костных пластинок поддерживали широкий ласт. Плечевой и тазовый пояса были слабо развиты. На теле имелось несколько плавников. Ихтиозавры были живородящими животными.
Наряду с ихтиозаврами жили плезиозавры. Появившиеся в среднем триасе, уже в
нижней юре достигли расцвета, в мелу они были обычны во всех морях. Они
подразделялись на две главные группы: длинношеих с маленькой головой (собственно
плезиозавры) и короткошеих с довольно массивной головой (плиозавры). Конечности
превратились в мощные ласты, ставшие главным органом плавания. Более примитивные
юрские плиозавры происходят, главным образом, с территории Европы. Плезиозавр из
нижней юры, достигал в длину 3 м. Эти животные еще часто выходили на берег для
отдыха. Плезиозавры не были столь ловкими в воде, как плиозавры. Этот недостаток
в известной мере восполнялся развитием длинной и очень гибкой шеи, с помощью
которой плезиозавры могли молниеносно схватывать добычу. Питались они
преимущественно рыбой и моллюсками.
В юрский период появляются новые роды ископаемых черепах, а в конце периода – и
современные черепахи.
Бесхвостые лягушкообразные земноводные обитали в пресных водоемах.
В юрских морях было очень много рыб: костистых, скатов, акул, хрящевых,
ганоидных. Они имели внутренний скелет из гибкой хрящевой ткани, пропитанной
солями кальция: плотный костный чешуйчатый покров, хорошо защищавший их от
врагов, и челюсти с крепкими зубами.
Из беспозвоночных в юрских морях водились аммониты, белемниты, морские лилии.
Однако в юрский период аммонитов насчитывалось гораздо меньше, чем в триасе.
Юрские аммониты отличаются от триасовых и по своему строению, за исключением
филоцераса, совершенно не изменившегося при переходе из триаса в юру. Отдельные
группы аммонитов сохранили до нашего времени перламутр. Одни животные обитали в
открытом море, другие – заселяли заливы и мелкие внутриконтинентальные моря.
Головоногие моллюски – белемниты – целыми стаями плавали в юрских морях.
Наряду с небольшими экземплярами были настоящие великаны – длиной до 3 м.
Остатки внутренних раковин белемнитов, известные под названием “чертовы пальцы”,
встречаются в отложениях юрского периода.
В морях юрского периода получили значительное развитие также двустворчатые
моллюски, особенно принадлежащие к семейству устриц. Они начинают образовывать
устричные банки. Значительные изменения претерпевают морские ежи, селившиеся на
рифах. Наряду с дожившими до наших дней круглыми формами жили двусторонне
симметричные неправильной формы ежи. Их тело было вытянуто в одном направлении.
Некоторые из них обладали челюстным аппаратом.
Юрские моря были относительно мелководными. Реки приносили в них мутную воду,
задерживая газообмен. Глубокие бухты наполнялись гниющими остатками и илом,
содержащим, большое количество сероводорода. Именно поэтому в подобных местах
хорошо сохранились остатки животных, занесенные морскими течениями или волнами.
Появляется много ракообразных: усоногие, десятиногие, листоногие раки,
пресноводные губки, среди насекомых – стрекозы, жуки, цикады, клопы.
С юрскими отложениями связаны месторождения угля, гипса, нефти, соли, никеля и кобальта.
И сменился мелом, и имел продолжительность около 56 млн лет.
География и климат
В юрский период суперконтинент Пангея начал раскалываться на два отдельных континента:
- северную часть, известную как Лавразия (которая в конечном итоге разделилась на Северную Америку и Евразию, открыв бассейны для Атлантического океана, и Мексиканского залива)
- южная часть - Гондвана - дрейфовала на восток (и витоге разделилась на Антарктику, Мадагаскар, Индию и Австралию, а ее западная часть, сформировала Африку, и Южную Америку).
Этот процесс разделения Пангеи наряду с более теплой глобальной температурой, позволил таким рептилиям, как динозавры диверсифицироваться и доминировать длительное время на Земле.
Жизнь растений
В эпоху мезозоя растения развивали способности вести наземный образ жизни и не ограничиваться только океанами. К началу юры, жизнь от мохообразных, низкорослых моховидных и печеночников, которые не имели сосудистой ткани и были ограничены влажными болотистыми участками.
Гинкговые деревья
Папоротники и гингковые, обладающие корнями и сосудистой тканью для транспортировки воды, и питательных веществ, а также размножающиеся споровым способом, были доминирующими растениями ранней юры. Во время юрского периода появился новый способ размножения растений. Голосеменные растения, такие как хвойные деревья, развили пыльцу, которая с помощью ветра распределяется на большие расстояния и опыляет женские шишки. Этот способ размножения позволил значительно увеличить численность голосеменных растений к концу юрского периода. Цветковые растения не эволюционировали до мелового периода.
Эра динозавров
Как было показано в фильме «Парк юрского периода», рептилии были доминирующей формой жизни животных в юрский период. Они преодолели эволюционные препятствия, которые ограничивали . У рептилий были сильные окостеневшие скелеты с передовыми мышечными системами для поддержки и передвижения тела. Некоторые из крупнейших животных, которые когда-либо жили, были динозаврами юрского периода. Рептилии также могли развивать амниотические яйца, которые инкубировались на суше.

завроподы
Завроподы (ящероногие динозавры) - травоядные четвероногие животные, с длинными шеями и тяжелыми хвостами. Многие завроподы такие, как брахиозавры, были огромными. Представители некоторых родов имели длину тела около 25 м, а вес колебался в пределах 50-100 тонн, что делает их крупнейшими наземными животными, которые когда-либо существовали на Земле. Их черепа были относительно маленькими, с ноздрями, высоко поднятыми к глазам. Такие маленькие черепа означали наличие очень маленьких мозгов. Несмотря на небольшие мозги, эта группа животных процветала в течение юрского периода и имела широкое географическое распределение. На всех континентах обнаружены окаменелости завропод, за исключением Антарктиды. Другие известные динозавры юры включают стегозавров и летающих птерозавров.
Карнозавры были одними из главных хищников мезозойской эры. Род аллозавров был одним из самых распространенных карнозавров в Северной Америке. Они похожи на более поздних тиранозавров, хотя проведенные исследования показывали, что у них мало общего. Аллозавры имели сильные задние конечности, тяжелые передние лапы и длинные челюсти.
Ранние млекопитающие

Аделобазилевс
Возможно, динозавры были доминирующими наземными животными, но не единственными представителями фауны. Ранние млекопитающие были в основном очень маленькими травоядными или насекомоядными животными, и не конкурировали с более крупными рептилиями. Аделобазилевс - хищный предок млекопитающих. Он имел особенное строение внутреннего уха и челюстей. Это животное появилось в конце триасового периода.
В августе 2011 года ученые из Китая объявили об открытии Юрамайи. Это крошечное животное середины юры вызвало волнение среди ученых, потому что это был явный предок плацентарных млекопитающих, свидетельствующий о том, что млекопитающие развивались намного раньше, чем считалось прежде.
Морская жизнь

Плезиозавр
Юрского периода также была весьма разнообразной. Крупнейшими морскими хищниками были плезиозавры. Эти плотоядные морские рептилии обычно имели широкие тела и длинные шеи с четырьмя конечностями в форме ласты.
Ихтиозавр - морская рептилия, был наиболее распространен в раннем юрском периоде. Поскольку некоторые окаменелости были найдены с более мелкими особями своего вида внутри тела, предполагается, что эти животные, возможно, были одними из первых, у кого была внутренняя беременность, и они рождали живых детенышей.
Головоногие также были широко распространены в юрский период и включали предков современных кальмаров. Среди самых красивых окаменелостей морской жизни можно выделить спиралевидные раковины аммонитов.



